

FY2011 Annual Report
Computational Neuroscience Unit
Professor Erik De Schutter

Abstract
We use computational, data-driven methods to study how neurons and microcircuits in the brain operate. We are interested in the interaction between fundamental properties like morphology or excitability and neural functions like information processing or learning. Most of our models concern the cerebellum as this brain structure has a relatively simple anatomy and the physiology of its main neurons has been studied extensively, allowing for detailed modeling at many different levels of complexity.
In FY2011 significant progress has been made on several research projects.
1. Staff
General services and neuroinformatics
- Ivan Raikov, Technical Staff
- Tsuyuki Nakabayashi, Research Administrator / Secretary:
Molecular modeling
- Gabriela Antunes, Researcher (till September 2011)
- Weiliang Chen, Researcher
- Iain Hepburn, Technical Staff
Cellular modeling
- Sungho Hong, Researcher
- Shiwei Huang, Researcher
- Yihwa Kim, Researcher (till January 2012)
- Haroon Anwar, Techmical Staff
- Hermina Nedelescu, Graduate Student
- Pascal Warnaar, Graduate Student
Network modeling
- Tom Close, Researcher (from June 2011)
- Fabio M.S. de Souza, Researcher (till September 2011)
- Daqing Guo, Researcher (from June 2011)
- Mario Negrello, Researcher
- Shyam Kumar Sudhakar, Graduate Student
2. Collaborations
Theme: Cerebellar physiology, multiple themes
- Type of collaboration: Scientific collaboration and graduate program
- Researchers:
- Professor M. Giugliano, University of Antwerp, Belgium
- Professor D. Snyders, University of Antwerp, Belgium
- Dr. Q. Robberecht, University of Antwerp, Belgium
- Dr. K. Tahon, University of Antwerp, Belgium
- Dr. K. Veys, University of Antwerp, Belgium
Theme: Spiking activity of monkey cerebellar neurons
- Type of collaboration: Scientific collaboration
- Researchers:
- Professor H.P. Thier, University of Tübingen, Germany
- Dr. A. Ignashchenkova, University of Tübingen, Germany
- Dr. M. Junker, University of Tübingen, Germany
- Dr. A. Schmigdlin, University of Tübingen, Germany
Theme: Molecular identification of cerebellar signaling pathways and cerebellar optogenetics
- Type of collaboration: Scientific collaboration
- Researchers:
- Professor G.J. Augustine, Korea Institute for Science and Technology (KIST), Korea
- Professor K. Tanaka, Korea Institute for Science and Technology (KIST), Korea
- Dr. J. Kim, Korea Institute for Science and Technology (KIST), Korea
Theme: Molecular modeling of diffusion in dendrites
- Type of collaboration: Scientific collaboration
- Researchers:
- Professor F. Santamaria, University Texas, San Antonio, United States of America
Theme: Ultrastructural morphology of Purkinje cells
- Type of collaboration: Scientific collaboration
- Researchers:
- Professor M. Ellisman, University California, San Diego, United States of America
- Professor M. Martone, University California, San Diego, United States of America
- Dr. L. Fong, University California, San Diego, United States of America
Theme: Modeling of effects of ethanol on the cerebellum
- Type of collaboration: Joint research
- Researchers:
- Professor C.F. Valenzuela, University of New Mexico, United States of America
Theme: Correlation of neurons linked to their excitability
- Type of collaboration: Scientific collaboration
- Researchers:
- Professor S.A. Prescott, University of Pittsburgh, United States of America
- Dr. S. Ratté, University of Pittsburgh, United States of America
Theme: Purkinje cell morphology and physiology, modeling
- Type of collaboration: Scientific collaboration
- Researchers:
- Professor M. Häusser, University College London, United Kingdom
- Dr. H. Cuntz, Goethe University Germany
- Dr. A. Roth, University College London, United Kingdom
- Dr. A. Watt, McGill University, Canada
3. Activities and Findings
3.1 Neuroinformatics standards
A Standard Language for Multiscale Modeling in Neuroscience
The growing number of large-scale neuronal network models has created a need for standards and guidelines to ease model sharing and facilitate the replication of results across different simulators. To foster community efforts towards such standards, the International Neuroinformatics Coordinating Facility (INCF) has formed its Multiscale Modeling program led by Prof. De Schutter, and has assembled a task force of simulator developers to propose a declarative computer language for descriptions of large-scale neuronal networks.
During the past year, we contributed to the efforts of the INCF Task Force on Multiscale Modeling towards releasing the first version of the NineML model description language, which is a computer language intended for descriptions of networks of integrate-and-fire neurons. The first version of the NineML specification and accompanying software has been released, and further developments of the language are ongoing.
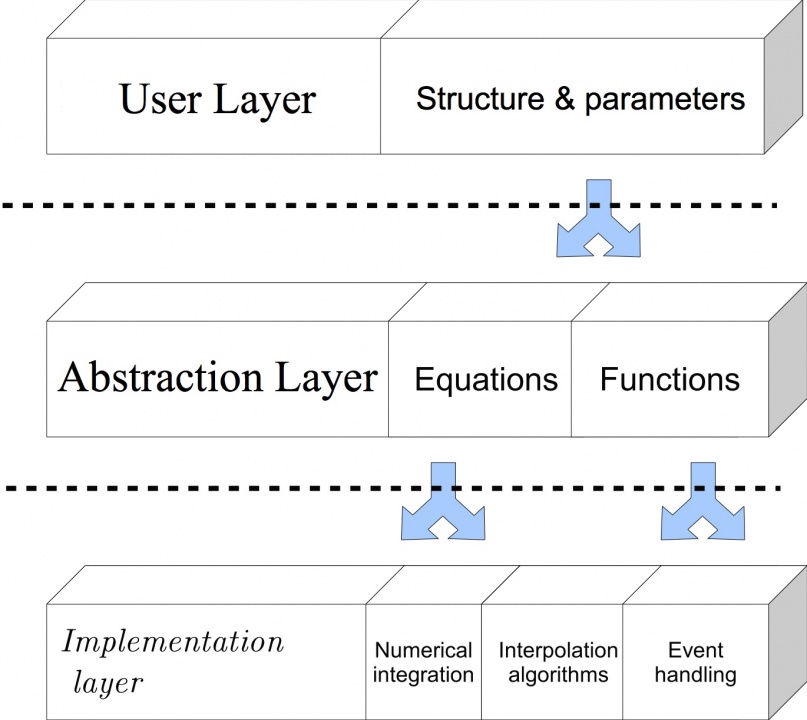 An important guiding principle to the design of NineML is the layer-oriented approach (Figure 1), which we proposed and developed as part of an earlier effort to develop a markup language for descriptions of neuronal ionic currents. A paper describing this approach appeared in PLoS Computational Biology (Raikov & De Schutter, 2012).
An important guiding principle to the design of NineML is the layer-oriented approach (Figure 1), which we proposed and developed as part of an earlier effort to develop a markup language for descriptions of neuronal ionic currents. A paper describing this approach appeared in PLoS Computational Biology (Raikov & De Schutter, 2012).Following the first release of NineML, we refocused our efforts to use the software infrastructure developed for NineML to develop a library for describing connectivity of neuronal networks based on anatomical geometric configurations of cells and layers in the cerebellum.
Figure 1: The layer-oriented approach
3.2 Molecular mechanisms of synaptic plasticity
STEPS software development
In FY2011 the STEPS development team released STEPS 1.3.0, a major update of the stochastic reaction-diffusion simulator. An important change in STEPS 1.3.0 is the implementation of new SSA kernel for the Tetexact solver. Previous versions of STEPS implemented the Gillespie Direct SSA method (Gillespie 1977) with Gibson and Bruck's optimization (Gibson & Bruck, 2000). By using a k-ary tree data structure, the implementation has an overall logarithmic complexity. In 1.3.0, we replace it by the new implementation of Composition and Rejection SSA (Slepoy et al., 2008). Via grouping propensities by their magnitudes and using rejection sampling, CR SSA achieves a constant time complexity, in both search and update of propensities. The new implementation of STEPS maintains the same accuracy as previous versions, yet is computationally more efficient.
In FY2011 the STEPS development team released STEPS 1.3.0, a major update of the stochastic reaction-diffusion simulator. An important change in STEPS 1.3.0 is the implementation of new SSA kernel for the Tetexact solver. Previous versions of STEPS implemented the Gillespie Direct SSA method (Gillespie 1977) with Gibson and Bruck's optimization (Gibson & Bruck, 2000). By using a k-ary tree data structure, the implementation has an overall logarithmic complexity. In 1.3.0, we replace it by the new implementation of Composition and Rejection SSA (Slepoy et al., 2008). Via grouping propensities by their magnitudes and using rejection sampling, CR SSA achieves a constant time complexity, in both search and update of propensities. The new implementation of STEPS maintains the same accuracy as previous versions, yet is computationally more efficient.
A comparison was made between STEPS 1.3.0 and other reaction-diffusion simulators, including MesoRD (Hattne et al., 2005) and Smoldyn (Andrews et al., 2010). With similar accuracy, STEPS 1.3.0 outperforms the other two by a significant margin (Hepburn et al., 2012). This is shown in Figure 2, where filled circles represent points where the simulation is calculated to be accurate and open circles show where simulation may be inaccurate. Results for an initial condition with a low number of molecules (A) is compared with that of a high number of molecules (B). Fo rSTEPS and MesoRD simulation runtimes at different number of subvolumes describing the same total mesh volumeare shown while for Smoldyn runtimes with varying number of simulation iterations due to a change in time-stepare shown. STEPS achieves high accuracy at faster runtimes than the other two simulators.
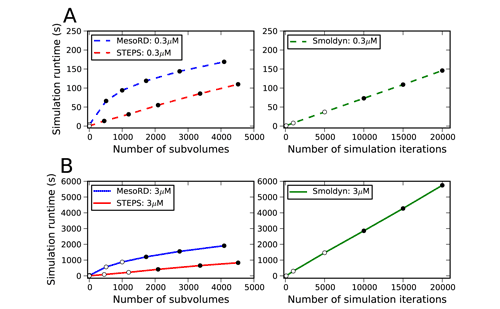
Figure 2: Efficiency of STEPS compared to MesoRD and Smoldyn.
3.3 Cellular mechanisms regulating firing and synaptic properties of neurons
Stochastic mechanisms in dendritic calcium spike generation
Neuronal activity is largely governed by molecular interactions taking place in and around complex cellular morphology. Intrinsic sources of noise in these interactions can have an important affect on neuronal signaling, though the extent and significance of this effect is not well understood for many neuronal systems. Such sources of noise include the stochastic interactions between diffusing and surface bound molecules, stochastic transitions between ion channel conformations and variability in aspects such as the cell morphology and molecular arrangements. Study of the influence of these factors is very important for our understanding of many systems, yet the complexity of such problems means that intuition alone is insufficient and computational studies of the represented systems become crucial.
We studied the stochastic behavior of dendritic calcium spikes in Purkinje neurons. To investigate the effect of the different sources of noise in this system we constructed a biologically detailed molecular model (Anwar et al. 2010) and ran simulations in STEPS (Hepburn et al., 2012), which supports stochastic and deterministic molecular simulations alongside accurate computation of the electrical behavior of the cellular region, all within complex 3D morphologies. Therefore we could run a series of stochastic, deterministic, and hybrid simulations and investigate the different sources of noise individually and together, and compare to deterministic solutions.
By comparing the sources of noise in this system in absolute and relative terms at different lengths of dendritic section we demonstrate the significance of the different contributing factors to stochasticity in the system, including calcium dynamics and ion channel clustering, on a variety of spatial scales.
Chloride dynamics in Purkinje cells: a computational and electrophysiological study
This study aims to investigate the physiological role of the uncoupled chloride current of glutamate transporters in Purkinje cells. Neuronal glutamate transporters take up synaptic glutamate to maintain a 106 concentration difference across the cell membrane, thus help maintain normal synaptic excitability. Purkinje cells express a unique class of glutamate transporters found predominantly on the spines of their dendritic arbor. Contrary to the conventional role of glutamate transporters, these Purkinje cell-specific glutamate transporters maintain normal synaptic excitability by ‘buffering’ synaptic glutamate. The binding of glutamate to these transporters also activate a thermodynamically uncoupled chloride current. The physiological role of this current is unknown.
A detailed model describing the synaptic activation of these glutamate transporters have been developed to explore the effects of the uncoupled chloride current and intracellular chloride accumulation on synaptic excitability. Simulation results of this model are being tested electrophysiologically.
Efficient Estimation of Phase Response Curves via Compressive Sensing
The phase response curve (PRC), relating the phase shift of an oscillator to external perturbation, is an important tool to study neurons and their population behavior. It can be experimentally estimated by measuring the phase changes caused by probe stimuli. These stimuli, usually short pulses or continuous noise, have a much wider frequency spectrum than that of neuronal dynamics. This makes the experimental data high dimensional while the number of data samples tends to be small. Current PRC estimation methods have not been optimized for efficiently discovering the relevant degrees of freedom from such data.
We propose a systematic and efficient approach based on a recently developed signal processing theory called Compressive Sensing (CS). CS is a framework for recovering sparsely constructed signals from undersampled data and is suitable for extracting information about the PRC from finite but high dimensional experimental measurements. We illustrate how the CS algorithm can be translated into an estimation scheme, and demonstrate that our CS method can produce good estimates of the PRCs with simulated and experimental data, especially when the data size is so small that simple approaches such as naive averaging fail. The tradeoffs between degrees of freedom versus goodness-of-fit were systematically analyzed, which help us to understand better what part of the data has the most predictive power (Hong et al. 2012).
In Figure 3 we compute PRCs from experimental data obtained from cerebellar Golgi cells with differennt levels of noisy current injection (indicated in the legends). We compare our CS method (A) with the commonly used WSTA (weighted spike-triggered average) method (B, Ota et al., 2009). While the 3 PRCs obtained with the CS method look very similar, as expected, those obtained with the WSTA method are much noiser and difficult to compare with each other.
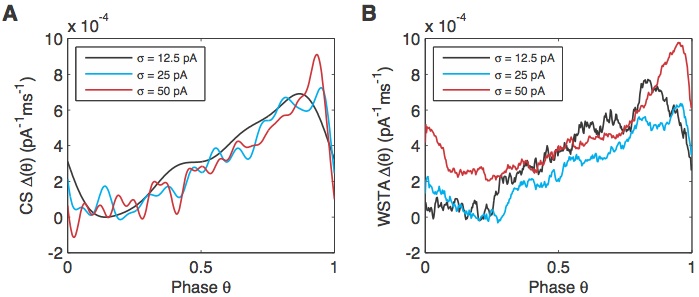
Figure 3: comparison of experimental PRCs obtained with our CS method (A) to those obtained with the WSTA method (B).
Our results illustrate that finite sizes of neuroscientific data in general compounded by large dimensionality can hamper studies of the neural code, and suggest that CS is a good tool for overcoming this challenge.
A Purkinje cell in the awake monkey produces multiple stable complex spike waveforms
Strong controversy exists about the role of complex spikes in cerebellar Purkinje cells. Complex spikes come in different shapes which are influenced by the state of the Purkinje cell as well by bursting activity in the climbing fiber input signal. The waveform therefore could harbor coding information, reflecting conditions in different parts of the olivo-cerebellar system. The shape of the complex spikes can influence intracellular calcium levels in the Purkinje cell, which are critically involved in plasticity or can influence the output to the cerebellar nuclei and thus encode additional information.
In this study complex spikes waveforms are described for the first time in the awake monkey obtained from the lab of Prof. P. Thier (University of Tübingen), based on 12 extracellular recordings, spanning more than an hour each. In total over 3000 complex spikes from 10 different Purkinje cells were analyzed and described in respect of their stable variations of shape. On average a single Purkinje cell shows 12±5 stable waveform shapes. Overall complex spikes can have 2 to 9 spikelets, within complex spikes of one Purkinje cell the range of spikelets varies from 2 till 4. The number of spikelets as well as onsets of the spikelets in different complex spikes stable waveforms were analyzed. We found a significant correlation between the presence of specific stable waveforms and the preceding inter complex spikes interval length in many recordings.
3.4 Information processing in the olivocerebellar system
Visualizing the developing Purkinje cell dendritic forest
Neuroanatomists have long studied at the level of single cells the morphological development of Purkinje cells with their remarkable sagittally-oriented planar dendritic trees. Extrapolating from these experiments, it has been proposed and generally accepted that Purkinje cell dendrites may be highly organized at the network level (Palkovitz et al., 1971). Given the technical difficulties in visualizing what has been called the “impenetrable neuronal forest”, much of this has been speculative. The prevalent Purkinje cell network model predicts that dendrites are parasagittally aligned in parallel rows with dendritic fields slightly offset, and overlapping across neighboring rows. The model also depicts a type of tiling arrangement of non-overlapping dendritic fields that belong to cells residing in the same row. Recent advances in transgenic mouse technology now allow us to investigate how Purkinje cell dendrites are organized at the network level, and to measure if their growth is coordinated within the developing cerebellum.
Using Cre-Lox Brainbow technology, we drive recombination of combinatorial XFPs in mouse cerebellar PCs. This enables us to spectrally resolve the dendritic arbors of hundreds of neighboring PCs from lobule V, at several postnatal (P) ages, including P7, 14, 30, and P ≥ 60 (adult) using confocal microscopy. Dendritic arbors from ~100 neighboring PCs were manually reconstructed from confocal image stacks at each developmental time point (Figure 4 and movie). The 3D reconstructed trees were then analyzed using Neurolucida Explorer and the TREES toolbox (Cuntz et al., 2011).

Figure 4: Brainbow stacks from cerebellum of a P30 mouse (A) and the reonstructed Purkinje cell forest (B).
Contrary to the prevalent Purkinje cell network model, our data prominently demonstrate that Purkinje cell dendritic arbors are not perfectly aligned in parallel rows; rather, individual dendrites slant bi-directionally away from a parallel row arrangement. Determining the fine structural architecture of the Purkinje cell network, and how it emerges developmentally is important for our understanding of cerebellar function.
Using software infrastructure built to support the NineML language, we have implemented a prototype framework for exploring hypotheses about the neuroanatomical structures and connectivity in the cerebellar cortex at various levels of granularity, based on experimental data and hypotheses from the scientific literature.
The tentative name of this framework is the NineML BREP (boundary representations). NineML BREP is an extension of NineML for constructive 3D boundary representation (Havemann, 2005) of neuroanatomical structures and connectivity at various levels of granularity (from coarse-resolution solids to fine meshes). NineML BREP provides the ability to specify geometric parameters for the instantiation of topological objects, such as coordinates for placement, or probability distributions for random placement of a group of identical objects; define categories of topological objects, such as stellate, basket and Golgi cells; define rules for connectivity between different categories of objects.
The core assumptions of the NineML BREP framework are: 1) connectivity rules are specified as probability distributions for overlapping volumes of objects of different categories; 2) synapse locations are randomly generated from the distribution associated with an overlapping volume 3) the volumes that represent dendritic trees have regions of uniform synaptic density.
As illustrated in Figure 5, the software framework currently being implemented consists of declarative and algorithmic components. The declarative components include languages for describing connectivity and neuronal and synaptic mechanisms, built as an extension to NineML. The algorithmic components are Python scripts and the PyNN program, which are used for interfacing to specific simulator platforms and for simulation control.
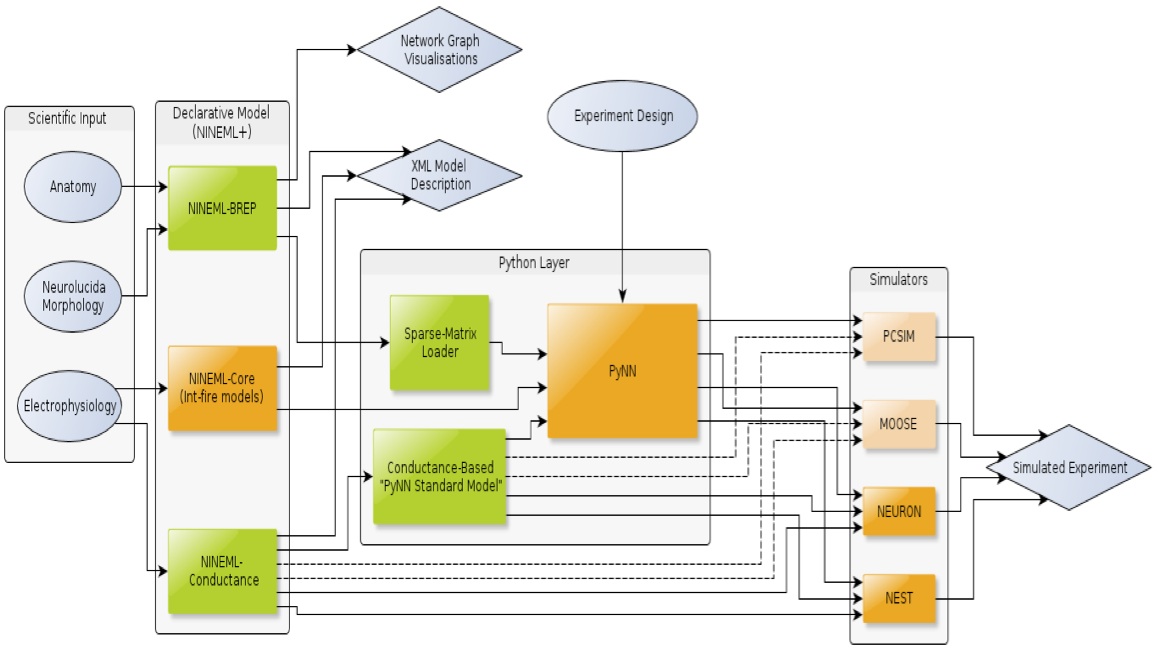
Figure 5: Conversion of an xml-based description of a network model of the cerebellum into code used by simulation software. NINEML-BREP is described here while NINEML-Conductance was described in Raikov and De Schutter (2012). The main target simulators are NEURON and NEST (full arrows).
Spread of inhibition through molecular layer gap junctions in the cerebellar cortex
The Purkinje cells of the cerebellar cortex receive inhibitory connections from molecular layer interneurons (basket and stellate cells). Basket cells synapse with the Purkinje cell soma forming a typical basket like structure (pinceau) thereby providing a powerful inhibition. Stellate cells synapse with Purkinje cell dendritic tree and forms many synapses with a single Purkinje cell. It is well known that interneurons of the molecular layer of cerebellar cortex are connected by electrical synapses. Here we replicated an optogenetic mapping experiment that quantified how molecular layer interneurons are spatially linked by gap junctions, thereby increasing the amount of inhibition reaching the target Purkinje cells (Kim et al. SfN 2011 Abstract #305.19).
We developed a 3D network model of the molecular layer of the cerebellar cortex. Basket and stellate cells were modeled as leaky integrate and fire neurons, tuned according to their firing properties. The interneurons were connected by gap junctions and GABAergic synapses and synaptically connected to Purkinje cells according to anatomical patterns and tuned to match the optogenetic data. To simulate the optical stimulus the experimentally measured footprints were used to estimate the number of interneurons activated. The network model predicts that Purkinje cells in the mouse receive direct input from 4-5 molecular layer interneurons but that up to 12 gap-junction connected interneurons can effectively inhibit the neuron.
Contributions of Purkinje cell pauses to saccadic eye movements
The saccadic eye movement resembles a slingshot in the onset, and reins, in the offset, in that it is the brisk release of a tense elastic muscle that allows a brisk movement initiation, while resuming the tension brings de-acceleration. Contrary to established belief, we provide compelling evidence that pauses of the cerebellar Purkinje neuron simple spikes, rather than bursts, are the reliable companions of saccades. This is intuitive, since disinhibition on the cerebellar nuclei is more likely to be functional than the burst, for while the increase of inhibition by a burst is gradual and moderate, the pause is absolute silence after noisy chatter.
This thesis is based on the analysis of cerebellar recordings from primates obtained from the lab of Prof. P. Thier (University of Tübingen), including single cell recordings and local field potentials as saccadic concomitants. Pauses are the most frequent concomitants of eye motion. For many cells more than 30% of the recorded saccades were linked to a pause, indicating that many cells participate in a considerable number of the saccades. They can be direction selective. Contrary to our expectations, the contribution of the complex spike for pauses has been scanty.
3.5 References cited
- Andrews S. S., Addy N. J., Brent R. & Arkin A. P. Detailed Simulations of Cell Biology with Smoldyn 2.1. PLoS Computational Biology 6:e1000705 (2010).
- Anwar H., Hong S. & De Schutter E. Controlling Ca2+-activated K+ channels with models of Ca2+ buffering in Purkinje cells. Cerebellum in press (2010).
- Cuntz H., Forstner F., Borst A. & Häusser M. The TREES Toolbox-Probing the Basis of Axonal and Dendritic Branching. Neuroinformatics 9: 91–96 (2011).
- Gibson M. A. & Bruck J. Efficient exact stochastic simulation of chemical systems with many species and many channels. Journal of Physical Chemistry 104:1876-1889 (2000).
- Gillespie D. T. Exact Stochastic Simulation of Coupled Chemical-Reactions. Journal of Physical Chemistry 1977, 81:2340-2361 (1977).
- Hattne J., Fange D. & Elf J. Stochastic reaction-diffusion simulation with MesoRD. Bioinformatics 21: 2923-2924 (2005).
- Havemann S. Generative Mesh Modeling, section 4.2 Boundary Representations and B-Rep Meshes. Ph.D. Dissertation, Technischen Universität Braunschweig (2005).
- Hepburn I., Wils S., Chen W. & De Schutter E. STEPS: Efficient simulation of stochastic reaction-diffusion models in realistic morphologies. BMC Systems Biology 6: 36 (2012).
- Hong S., Robberechts Q. & De Schutter E. Efficient estimation of phase response curves via compressive sensing.
- Palkovits M., Magyar P. & Szentágothai J. Quantitative histological analysis of the cerebellar cortex in the cat. I. Number and arrangement in space of the Purkinje cells. Brain research 32: 1–13 (1971).
- Raikov I. & De Schutter E. The layer-oriented approach to biological modeling languages. PLoS Computational Biology 8: e1002521 (2012).
- Slepoy A., Thompson A. P. & Plimpton S.J. A constant-time kinetic Monte Carlo algorithm for simulation of large biochemical reaction networks. Journal of Chemical Physics 128: 205101 (2008).
4. Publications
4.1 Journals
- Botta, P., Simoes de Souza, F. M., Sangrey, T., De Schutter, E. & Valenzuela, C. F. Excitation of Rat Cerebellar Golgi Cells by Ethanol: Further Characterization of the Mechanism. Alcoholism: Clinical and Experimental Research 36, 616-624, doi:10.1111/j.1530-0277.2011.01658.x (2011).
- Hepburn, I., Wils, S., Chen, W. & De Schutter, E. STEPS: Efficient simulation of stochastic reaction-diffusion models in realistic morphologies. BMC Systems Biology 6, 36 (2012).
- Hong, S., Ratte, S., Prescott, S. A. & De Schutter, E. Single neuron firing properties impact correlation-based population coding. Journal of Neuroscience 32, 1413-1428, doi:32/4/1413 [pii]10.1523/JNEUROSCI.3735-11.2012 (2012).
- Kennedy, D. N., Ascoli, G. A. & De Schutter, E. Next steps in data publishing. Neuroinformatics 9, 317-320, doi:10.1007/s12021-011-9131-0 (2011).
- Le Franc, Y., Giugliano, M. & De Schutter, E. The Belgian node: a community platform toward the future. Acta Neurologica Belgica 11 (2011).
- Raikov, I. & De Schutter, E. The Promise and Shortcomings of XML as an Interchange Format for Computational Models of Biology. Neuroinformatics 10, 1-3, doi:10.1007/s12021-011-9136-8 (2012).
- Santamaria, F., Wils, S., De Schutter, E. & Augustine, G. J. The diffusional properties of dendrites depend on the density of dendritic spines. European Journal of Neuroscience 34, 561-568, doi:10.1111/j.1460-9568.2011.07785.x (2011).
- Steuber, V., Schultheiss, N., Silver, A., De Schutter, E. & Jaeger, D. Determinants of synaptic integration and heterogeneity in rebound firing explored with date-driven models of deep cerebellar nuclei cells. Journal of Computational Neuroscience 30, 633-658, doi:10.1007/s10827-010-0282-z (2011).
- Tahon, K., Wijnants, M. & De Schutter, E. The RAT-ROTADRUM: A reaction time task depending on a continuous stream of tactile sensory information to the rat. Journal of Neuroscience Methods 200, 153-163, doi:S0165-0270(11)00385-2 [pii]10.1016/j.jneumeth.2011.06.031 (2011).
- Tahon, K., Wijnants, M., De Schutter, E. & Maex, R. Current source density correlates of cerebellar Golgi and Purkinje cell responses to tactile input. Journal of Neurophysiology 105, 1327-1341, doi:jn.00317.2010 [pii]10.1152/jn.00317.2010 (2011).
- Uusisaari, M. & De Schutter, E. The mysterious microcircuitry of the cerebellar nuclei. Journal of Physiology (London) 589, 3441-3457, doi:jphysiol.2010.201582 [pii]10.1113/jphysiol.2010.201582 (2011).
4.2 Books and other one-time publications
- Tahon, K. Cerebellar processing of tactile input. PhD thesis, University of Antwerp, Belgium (2011).
4.3 Oral Presentations
-
De Schutter, E. Stochastic modeling of signaling pathways involved in induction of cerebellar long-term depression, Experimental Biology 2011, Washington, DC, USA, April 9-13, 2011, 2011.
-
De Schutter, E. Describing anatomical connectivity in modeling of realistic networks, Molecular Neuroanatomy Workshop, Allen Institute for Brain Science, Seattle, WA, USA, May 25-29, 2011.
-
Hong, S. Single neuron firing properties impact correlation-based population coding, The 6th APCTP-KAIST School for Brain Dynamics: Young Computational Neuroscientist Workshop, Daejeon, South Korea, December 5, 2011, 2011.
-
Raikov, I. Workflow for Network Modeling with NineML, Computational Neuroscience Workshops, Stockholm, Sweden, July 27, 2011, 2011.
4.4 Poster Presentations
-
Antunes, G. & De Schutter, E. Stochasticity of the induction of cerebellar long-term depression: a signaling pathway modeling study, 8th IBRO World Conference of Neuroscience, Florence, Italy, July 9-13, 2011, 2011.
-
Anwar, H. & De Schutter, E. Compensating the effect of dendritic diameters on calcium transients: a modeling study, Computational Neuroscience, Stockholm, Sweden, July 23-28, 2011, 2011.
-
Anwar, H., Van Geit, W., Hong, S. & De Schutter, E. Constructing a biophysically detailed model of Purkinje cell dendrites, 41st Annual Meeting Society for Neuroscience, Washington DC, USA, November 12-16, 2011, 2011.
-
Chen, W., Hepburn, I. & De Schutter, E. Improving performance of the STochastic Engine for Pathway Simulation (STEPS), Computational Neuroscience, Stockholm, Sweden, July 23-28, 2011, 2011.
-
Gorchetchnikov, A., Cannon, R., Clewley, R., Cornelis, H., Davison, A., De Schutter, E., Djurfeldt, M., Gleeson, P., Hil, l. S., Hines, M., Kriener, B., Le Franc, Y., C.C., L., Morrison, A., Muller, E., Plesser, H. E., Raikov, I., Ray, S., Schwabe, L. & Szatmary, B. NineML: declarative, mathematically-explicit descriptions of spiking neuronal networks, Neuroinformatics 2011, Boston, Massachusetts, USA, September 4-6, 2011, 2011.
-
Hepburn, I., Chen, W. & De Schutter, E. SBML and MathML support for the stochastic reaction-diffusion simulator STEPS, Computational Neuroscience, Stockholm, Sweden, July 23-28, 2011, 2011.
-
Hong, S. & De Schutter, E. Efficient estimation of phase response curves via compressive sensing, Computational Neuroscience, Stockholm, Sweden, July 23-28, 2011, 2011.
-
Hong, S., Ratté, S., Prescott, S. A. & De Schutter, E. Single neuron firing properties impact correlation-based population coding, 41st Annual Meeting Society for Neuroscience, Washington DC, USA, November 12-16, 2011, 2011.
-
Huang, S. & De Schutter, E. The effect of glutamate-gated chloride current on the excitability of a Purkinje cell: a modeling study, Computational Neuroscience, Stockholm, Sweden, July 23-28, 2011, 2011.
-
Huang, S. & De Schutter, E. A computational study of chloride metabolism in Purkinje neuron, 41st Annual Meeting Society for Neuroscience, Washington DC, USA, November 12-16, 2011, 2011.
-
Kim, Y. & De Schutter, E. Repulsion between branches might be a key factor in generating cell type-specific dendritic bifurcation geometry, 41st Annual Meeting Society for Neuroscience, Washington DC, USA, November 12-16, 2011, 2011.
-
Negrello, M., Raikov, I. & De Schutter, E. Boundary representation of neural architecture and connectivity, Computational Neuroscience, Stockholm, Sweden, July 23-28, 2011, 2011.
-
Raikov, I., Cannon, R., Clewley, R., Cornelis, H., Davison, A., De Schutter, E., Djurfeldt, M., Gleeson, P., Gorchetchnikov, A., Plesser, H. E., Hil, l. S., Hines, M., Kriener, B., Le Franc, Y., C.C., L., Morrison, A., Muller, E., Ray, S., Schwabe, L. & Szatmary, B. NineML: the network interchange for neuroscience modeling language, Computational Neuroscience, Stockholm, Sweden, July 23-28, 2011, 2011.
-
Song, Z., Postma, M., Chen, W., Coca, D., Billings, S. A., Hardie, R. C., Juusola, M. & De Schutter, E. Does calcium diffusional global feedback leads to slow light adaptation in Drosophila photoreceptors? - A 3D biophysical modelling approach, Computational Neuroscience, Stockholm, Sweden, July 23-28, 2011, 2011
5. Intellectual Property Rights and Other Specific Achievements
Nothing to report
6. Meetings and Events
6.1 Summer courses
Okinawa Computational Neuroscience Course 2011
- Date: June 13 – June 30, 2011
- Venue: Seaside House, OIST
- Co-organizers : K. Doya, K. Stiefel, J. Wickens, OIST
- Co-sponsors : Japanese Neural Network Society
- Speakers:
- Arbuthnott, Gordon, OIST, Japan
- Brenner, Sydney, OIST, Japan
- De Schutter, Erik, OIST, Japan
- Doya, Kenji, OIST, Japan
-
Ermentrout, Bard, University of Pittsburgh, USA
-
Haruno, Masahiko, Tamagawa University, Japan
-
Kuhn, Bernd, OIST, Japan
-
Li, Zhaoping, University College London, UK
-
McCormick, David, Yale University, USA
-
Mongillo, Gianluigi, Université Paris Descartes, France
-
Prinz, Astrid, Emory University, USA
-
Seung, Sebastian, MIT, USA
-
Stevens, Chuck, Salk Institute, USA
-
Stiefel, Klaus, OIST, Japan
-
Toyoizumi, Taro, RIKEN BSI, Japan
-
Vaadia, Eilon, Hebrew University, Israel
-
Yu, Angela, University of Calformia San Diego, USA
- Date: October 10 - 21, 2011
- Venue: Seaside House, OIST
- Co-sponsor: The Allen Institute for Brain Science
- Co-organizers : K. Doya, OIST, John Hohmann, Terri Gilbert, AIBS
- Speakers:
- Brenner, Sydney, OIST, Japan
- Dang, Chin, AIBS, USA
- De Schutter, Erik, OIST, Japan
- Doya, Kenji, OIST, Japan
- Ellisman, Mark, UCSD, USA
-
Gilbert, Terri, AIBS, USA
-
Hohmann, John, AIBS, USA
-
Hof, Patrick, Mount Sinai School of Medicine, USA
-
Jones, Allan, AIBS, USA
-
Morris, John, AIBS, USA
-
Pules, Luis, University of Murcia, Spain
-
Watson, Charles, Curtin University of Technology, Perth, Australia
6.2 Seminars
Toward a global infrastructure for neuroscience data sharing and integration
- Date: October 121, 2011
- Venue: Campus, OIST
- Speaker: Sean Hill, INCF, Sweden
Role of striatal activity in Parkinson's disease related oscillations in the basal ganglia
- Date: March 5, 2012
- Venue: Campus, OIST
- Speaker: Arvind Kumar from Bernstein Centre, University of Freiburg, Germany
7. Other
Nothing to report.